Сбор и первичная обработка
Методы
оценки численности популяций
Количественные
учёты - один из важнейших компонентов
экологических исследований, направленных
на познание закономерностей
пространственного распределения видов,
динамики их популяций, функционирования и
организации многовидовых сообществ.
Сведения о численности отдельных видов,
кроме того, имеют практическое значение,
особенно в сфере изучения охотничье-промысловых
животных, переносчиков заболеваний,
вредителей сельского хозяйства, а также
редких и исчезающих видов [Кашкаров, 1933 и мн.
др.]. Такие данные используются и при
изучении фенотипической изменчивости
организмов [Тимофеев-Ресовский и др.,
1969].
Методы
учёта различных таксономических и
экологических групп организмов настолько
разнообразны, что нет возможности охватить
их в кратком обзоре. Однако вопрос этот
чрезвычайно важен, так как универсальных
приёмов не существует, а ошибку в
проведении учётов значительно труднее
исправить, чем ошибку в их последующей
обработке. Поэтому, прежде чем приступать к
количественным учётам, лучше потратить
время на поиск и прочтение специальной
литературы [Бей-Биенко, 1932; Методы …, 1952;
Любищев, 1958; Организация …, 1963; Василевич,
1969; Свиридов, Водопьянов, 1977; Количественные
…, 1987; Равкин, Челинцев, 1990; Пшеницына и др.,
1993 и др.]. Здесь мы рассмотрим лишь в
первом приближении наиболее известные
методики.
Для
выявления обилия отдельных видов и их
соотношений в сообществах нередко пригоден
метод учётных
площадок. Учётная площадка, в самом общем
виде, представляет собой ограниченный тем
или иным способом участок земной
поверхности, на котором производится
подсчёт организмов. Размеры учётных
площадок зависят от объекта исследования и
могут колебаться в пределах от нескольких см2
(при изучении численности мелких
членистоногих или мелких растений) до
десятков м2 и
более (при лесотипологических работах или
учёте крупных млекопитающих). Для получения
статистически значимых результатов в
пределах каждого местообитания таких
площадок закладывается несколько.
Метод
трансектов (лат. trans - через и sectio -
разрезание) применяется для учёта хорошо
заметных организмов (сосудистые растения,
птицы, дневные бабочки, стрекозы, шмели и т.п.).
Он состоит в том, что учётчик движется по
маршруту известной протяженности и
регистрирует всех представителей
исследуемой группы организмов в пределах
полосы определенной ширины. Полученные
данные о численности видов в пределах
фактически обследованной площади в
дальнейшем, для сопоставимости,
пересчитываются на определённую единицу
площади (число экземпляров /м2,
/га или /км2). В геоботанических работах
ширина трансекта может быть сведена до
одной линии [Быков, 1967]. Для подсчета
пресмыкающихся и земноводных обычно
используют трансект шириной 3 метра (1.5
метра справа и слева от учётчика) [Динесман,
Калецкая, 1952]. В условиях разреженного
травянистого покрова этот метод может
использоваться и для подсчёта мелких
членистоногих - цикадовых, клопов, мелких
двукрылых и перепончатокрылых, жуков,
клещей, пауков и т. п., но тогда ширина
трансекта составляет 0.25 - 1 метр [Второв, 1968;
Злотин, 1975 и др.].
Дневных
бабочек учитывают обычно на полосе шириной
3 метра [Yamomoto, 1975; Pollard, 1977; Сергеев, Дубатолов,
1988; Дубатолов, Костерин, 1999 и др.]. Однако
более эффективно рассчитывать ширину
трансекта для каждого вида отдельно по
удвоенной средней дальности его
обнаружения [Малков, 1994; Малков, Малков, 1999;
Бондаренко, 1999, 2003]. Для вычисления обилия в
таком случае можно использовать формулу:
![]() ,
(2.1.)
,
(2.1.)
где xi - обилие i
вида в пересчете на 1 гектар, ni
- число зарегистрированных особей i
вида, k i -
средняя дальность обнаружения i вида
в метрах, L - длина
маршрута в километрах.
При
учёте птиц особи регистрируются как путём
визуального наблюдения, так и на слух,
поэтому их лучше разделять на три группы по
дальности фактического обнаружения: 1)
птицы, обнаруженные близко - до 25 м от учётчика; 2) недалеко - в 25 - 100 м от учётчика; 3) далеко - от 100 до 300 м от учётчика.
Удобнее
использовать формулу:
![]() ,
(2.2.)
,
(2.2.)
где
xi - обилие i
вида в пересчете на 1 км2,
б - число особей i вида, замеченных близко, н
- недалеко, д - далеко,
L - длина маршрута в
километрах [Равкин, 1967].
При
изучении насекомых, обитателей травяного
яруса, нередко применяются различные
модификации метода кошения
энтомологическим сачком. Наиболее
известная его разновидность - учёт на время.
Для этого в исследуемом местообитании
насекомые отлавливаются стандартным
сачком (диаметром 40 см)
в течение определенного промежутка времени
с последующим пересчетом на 1 час [Бей-Биенко,
1932; Сергеев, 1986; Пшеницына и др., 1993].
Скрытно
живущих и ночных животных обычно
приходится отлавливать в разнообразные ловушки.
В начало
страницы
Техника
измерений и округлений
На первом
этапе изучения морфологии организмов
неизбежно встает вопрос о том, что и как
измерять, чтобы наилучшим образом отразить
специфику объекта исследования. Совершенно
очевидно, что на этот вопрос нет и не может
быть универсального ответа, однако по
некоторым, наиболее хорошо изученным
таксономическим группам животных
морфологические промеры стандартизированы.
Так, у мелких млекопитающих измеряют длину
тела, хвоста, уха, длину ступни задней ноги,
а также определенные параметры черепа [Бобринский
и др., 1965; Юдин, 1971; Попов, 1977], у птиц -
длину крыла, хвоста, клюва и плюсны [Иванов,
Штегман, 1964] и т.п. Перечисленные признаки
важны для определения, поэтому их наносят
на этикетки, которые хранятся вместе с
коллекционными экземплярами. Для решения
более специальных задач, например, при
выяснении возрастной, половой,
географической изменчивости таксона имеет
смысл делать все возможные измерения и уже
в ходе последующего анализа выбирать
наиболее информативные признаки. При
описании собранного материала в научных
статьях, дипломных и курсовых работах
всегда необходимо подробно оговаривать,
каким образом проводили тот или иной замер,
допустим: "ширина крыла - расстояние от
кистевого сустава до вершины первого
второстепенного махового пера".
При
изучении морфологии организмов вовсе
необязательно делать упор на достижение
максимально возможной точности, а нужно
исходить лишь из соображений
целесообразности. Классики числовой
таксономии Дж. Симпсон и Э. Роэ [1939] показали
(цит. по: [Майр и др., 1956]), что, если в руках
исследователя имеется относительно
большая выборка, в качестве единицы
измерения следует принимать 1/20
разницы между самым крупным и самым мелким
экземпляром в выборке. Например, если
значения вариант укладываются в ряд от 10 до
12 мм, измерения следует проводить с
точностью до 0.1 мм ((12 –
10) / 20 = 0.1); если от 40 до 50 мм
- с точностью до 0.5 мм ((50 –40)
/ 20 = 0.5); если же пределами измерений служат 70
и 90 мм, то измерения следует вести в
целых миллиметрах и т.п.
В ходе вычисления обобщающих статистических показателей постоянно приходится сталкиваться с необходимостью округлений. Существуют разнообразные способы округления и, в принципе, можно использовать любой из них, важно лишь на протяжении всего хода работ следовать одному и тому же правилу. Обычно поступают следующим образом: если за последней сохраняемой цифрой следуют цифры <5, то они отбрасываются, если ³5, то последняя цифра увеличивается на единицу.
В начало
страницы
Отношения
и индексы
Абсолютные
размеры тела, как правило, весьма изменчивы
и зависят от возраста, условий
существования и т.п., однако их отношения
могут оставаться сравнительно постоянными.
Для выявления относительных размеров (индексов)
различных частей тела можно пользоваться
формулой:
![]() или
или
![]() (2.3.),
(2.3.),
где Ix - индекс меньшего по величине
признака, Iy -
индекс большего по величине признака, x
– значение меньшего признака, y
– значение большего признака. В тех
случаях когда признаки x и y примерно
равны по величине, возможна ситуация, при
которой отдельные значения признака y
могут оказаться меньшими.
В
качестве примера приведем процедуру
вычисления относительной длины хвоста, по
двум видам кротов: обыкновенному Talpa
europea L. и сибирскому Asioscalops
altaica Nikolsky (табл. 1). Определяем индекс
относительной длины хвоста:
для
обыкновенного крота
![]() ,
,
для
сибирского крота
![]() .
.
Таблица
1
Средние величины некоторых морфологических параметров кротов
T.
europea (I)
и A. altaica (II), мм
(по данным Б. С. Юдина [1971])
| Признак | I | II |
| Длина тела с головой | 133.6 | 153.8 |
| Длина хвоста | 25.2 | 27.0 |
Сравнивая
полученные результаты, приходим к выводу,
что обыкновенный крот, имея меньшие
абсолютные размеры, отличается бόльшей
относительной длиной хвоста.
Отношения
широко используются не только для
сравнительных целей, но и для определения
слабо различимых видов. Так, например, два
вида саранчовых - итальянский прус Calliptamus italicus L. и светлокрылый прус C. abbreviatus Ikonn.,
внешне очень похожи и их идентификация
могла бы представлять определенные
трудности, однако оказалось, что они без
проблем отличаются по отношению длины
надкрылий к длине заднего бедра [Ванькова,
1998].
Как
уже отмечалось, подвергая варьирующий
признак статистическому анализу, мы, в
сущности, отвлекаемся от конкретного
содержания и оперируем лишь абстрактными
значениями. При этом смысл анализа сводится
к выявлению общих свойств генеральной
совокупности, отраженных в конкретной
выборке. В связи с этим уже на самых ранних
этапах статистической обработки может
оказаться полезной группировка первичных
данных. Группировка считается успешной,
если удовлетворяет следующим требованиям [Тьюки,
1981].
1.
В ходе группировки не теряется важная
информация.
2.
В результате группировки первичные
данные принимают более компактную форму,
что в свою очередь облегчает восприятие и
последующие вычисления.
Во многих случаях этим положениям соответствует метод вариационных рядов, показывающий, каким образом числовые значения признака связаны с их повторяемостью в данной выборке [Плохинский, 1970; Лакин, 1990 и др.]. Вариационные ряды подразделяются на интервальные и безынтервальные. Безынтервальный вариационный ряд целесообразно строить в тех случаях, когда разница между минимальным и максимальным значением вариантов невелика, особенно, если признак варьирует дискретно. Последовательность его построения рассмотрим на примере данных о числе глазков на 40 клубнях картофеля. Результаты подсчёта оказались следующие: 7, 9, 5, 11, 9, 7, 8, 5, 8, 7, 7, 10, 6, 7, 5, 7, 8, 6, 5, 7, 7, 6, 5, 6, 4, 5, 5, 10, 5, 8, 7, 6, 7, 5, 7, 7, 6, 5, 6, 6. Находим минимальное и максимальное значение признака (xmin = 4, xmax = 11). Записываем все возможные, в пределах минимального и максимального, значения, и указываем, как часто принимает ту или иную величину анализируемый признак x. В общем случае частоты признака обозначаются буквой f (англ. frequency). Для их подсчета можно пользоваться условными обозначениями:
. = 1; . . = 2; : . = 3; : : = 4; I : = 5; Г: = 6; ГI = 7; [] = 8; [/] = 9; [x] = 10; [x] . = 11; [x] : = 12 и т.д.
Вариационный
ряд числа глазков на клубнях картофеля (n = 40)
| Значение признака x | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 |
| Символы частот признака fx | . | [x] | [] | [x] : | : : | . . | . . | . |
| Частота признака fx | 1 | 10 | 8 | 12 | 4 | 2 | 2 | 1 |
Полученный
вариационный ряд полностью соответствует
требованиям, предъявляемым к группировке
данных, поскольку первоначальный материал
стал более «обозримым», и в результате
простейших преобразований мы сделали явной
ранее завуалированную информацию. В
частности, хорошо видно, что большая часть
значений признака сгруппирована в пределах
5-7 глазков, тогда как крайние значения
вариант встречаются относительно редко.
Процедуру
построения интервального вариационного
ряда разберём на материале, собранном
студентами Г-АГУ. Измерение длины
предплечья у 53 особей летучей мыши вида
большой трубконос Murina
leucogaster Milne-Edwards без учета половой
принадлежности с точностью до 0.1 мм дало следующие результаты: 39.5, 40.4,
38.6, 39.4, 39.5, 40.0, 38.6, 36.0, 39.0, 40.4, 39.5, 42.5, 41.0, 39.6, 41.8,
39.5, 40.0, 39.7, 40.0, 37.5, 42.0, 38.7, 41.5, 40.0, 42.0, 40.0, 41.5, 40.5,
37.6, 40.0, 39.5, 40.7, 40.0, 41.7, 40.0, 41.5, 41.3, 41.2, 39.0, 40.0, 40.0,
42.0, 39.3, 40.5, 39.0, 40.5, 39.5, 40.0, 39.8, 42.0, 41.2, 40.0, 39.5.
Отыскиваем
минимальное и максимальное значение
признака (xmin=36.0,
xmax=42.5). Затем
находим величину классового интервала (l)
по формуле:
![]() ,
(2.4.)
,
(2.4.)
где
k - приблизительное число классов, на
которое следует разбить вариацию признака.
Значения k приведены
ниже (табл. 2). Как видно из таблицы, более
представительные по объёму выборки следует
разбивать на большее число классов.
Последнее требование лишь не значительно
сказывается на затратах времени, но зато
сводит к минимуму потерю информации.
Таблица
2
| Число наблюдений n | Число классов K |
| 20-30 | 5 |
| 30-100 | 7 |
| 100-1000 | 10 и более |
Подставляя
значения в формулу (2.4.), получаем величину
классового интервала (l=1.1 мм). Принимаем
минимальное значение длины предплечья
большого трубконоса (xmin=36.0) за середину первого
классового интервала. В результате
получаем следующие интервалы 35.5 - 36.6 - 37.7 - 38.8
- 39.9 - 41.0 - 42.1 - 43.2. После уменьшения верхних
границ классов на точность измерений (0.1 мм.)
и распределения значений вариантов по
классам получаем вариационный ряд (табл. 3).
Таблица
3
Вариационный
ряд длины предплечья большого трубконоса (n=53)
| Классовые интервалы x | 35.5-36.5 | 36.6-37.6 | 37.7-38.7 | 38.8-39.8 | 39.9-40.9 | 41.0-42.0 | 42.1-43.1 |
| Центральная величина классового интервала | 36.0 | 37.1 | 38.2 | 39.3 | 40.4 | 41.5 | 42.6 |
| Частота fx | 1 | 2 | 3 | 15 | 18 | 13 | 1 |
Для вычисления статистических параметров интервальный вариационный ряд трансформируют в безынтервальный. При этом середину классового интервала принимают за величину, характеризующую все варианты входящие в этот класс. Середину каждого классового интервала находят отношением суммы значений нижней и верхней границы интервала к 2. Отсюда центральная величина первого классового интервала равна 35.5+36.5/2=36.0, второго - 36.6+37.6/2=37.1, третьего - 37.7+38.7=38.2 и т.д.
Графическое отображение данных
График
(диаграмма) - одно из наиболее эффективных
средств представления данных. Правильно
построенный график может служить мощным
инструментом как для иллюстрации материала,
так и для формирования предварительных
гипотез. Неслучайно графики стали почти
обязательным элементом дипломных и
курсовых работ, многих научных публикаций,
основанных на количественных данных. Более
широкое распространение получили
двумерные графики (2 М), в том числе с
добавочной осью ординат, поскольку с них
удобнее считывать информацию. Из семейства
двумерных графиков наиболее известны
точечные диаграммы рассеяния, линейные
графики и столбчатые гистограммы.
Точечные диаграммы рассеяния строят, когда на графике необходимо сохранить информацию о каждом варианте анализируемой группы данных. В частности, по ним можно установить, существует ли между определенными признаками зависимость или они не связаны друг с другом. Кроме того, точечные диаграммы позволяют быстро определить, перекрывается ли один и тот же признак у представителей разных выборок. Пример такого графика приведен на рисунке 1, где отображено соотношение трёх признаков, проанализированных на экспонатах коллекции черепов алтайской популяции бурого медведя (зоомузей Г-АГУ). Из графика видно, что длина правого верхнего клыка достаточно равномерно возрастает при увеличении общей длины черепа, тогда как ширина мозговой капсулы значительно в меньшей степени зависима от этого показателя.
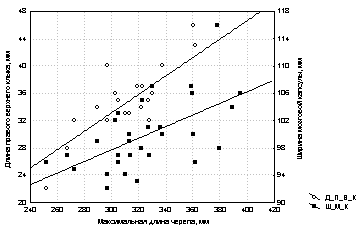
Рис.
1. Точечная
диаграмма, показывающая взаимосвязь между
некоторыми краниологическими признаками у
представителей алтайской популяции бурого
медведя.
На
линейных графиках отдельные точки
данных соединены линией. Обычно графики
такого типа используются, когда изучается
определенная последовательность значений,
причем порядок этих значений является
одним из существенных признаков. Линейные
графики могут быть очень полезными для
иллюстрации изменения какого-либо явления
во времени и потому они широко применяются
при изучении суточной, сезонной и вековой
динамики численности организмов, для
изучения процессов онтогенеза и т.п.. В
случае, если значения одной или нескольких
переменных изменяются на порядок и более,
график предпочтительнее строить в
логарифмическом масштабе [Уильямсон,
1975] (сравните
рис. 2 А и 2 Б).
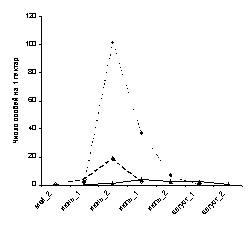
2
А
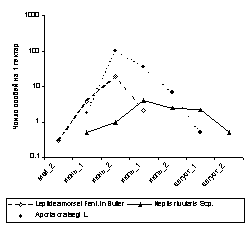
2
Б
Рис.
2. Линейные графики
динамики лёта некоторых дневных бабочек в
среднегорных березово-еловых лесах Северо-Восточного
Алтая. А - график построен в обычном
масштабе, Б - график построен в
логарифмическом масштабе.
Гистограмма
(термин
впервые использован Пирсоном в 1895 году)
представляет собой графическое
отображение вариационного ряда. На
гистограмме по оси абсцисс откладываются
границы классовых интервалов, а по оси
ординат – частоты. Затем для каждого класса
рисуется столбец, высота которого
пропорциональна частоте класса. Основное
назначение гистограмм, кроме
иллюстративной функции, - выявление
соответствия частот полученного в ходе
исследования выборки вариационного ряда
нормальному распределению. На рисунке 3
приведена гистограмма вариационного ряда,
построенного по промерам 53 особей большого
трубконоса (табл. 3).
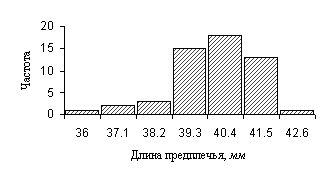
Рис.
3. Гистограмма,
иллюстрирующая изменчивость длины
предплечья у большого трубконоса на
Северном Алтае (n=53), мм.