Территория Юго-Восточного Алтая крайне интересна как с биогеографической (здесь проходит ряд важных фаунистических рубежей), так и экологической точек зрения. Последнее определяется, в первую очередь, своеобразием криоаридных экосистем и их относительно слабой нарушенностью. В некоторых физико-географических провинциях Алтая пространственно-типологическая организация населения мелких млекопитающих [21; 7] , птиц [14; 16] др., и дневных бабочек [10] изучена сравнительно полно. Для бабочек Юго-Восточного Алтая проведен зоогеографический анализ фауны, опубликован список видов, даны количественные оценки населения и, в частности, редких видов [8; 11; 9; 3; 22; 4; 5; ]. Однако обобщающей работы по сообществам дневных чешуекрылых этого региона Алтая до сих пор нет. Целью настоящей публикации является выявление пространственной структуры сообществ, классификация населения и факторов среды, определяющих ее характер.
Материал и методика
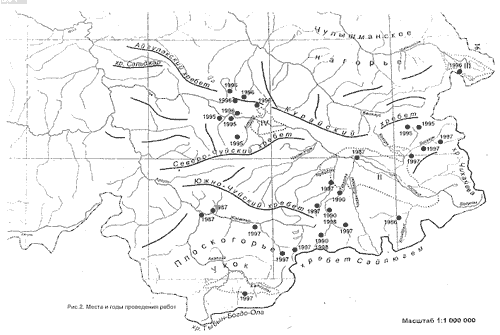
Учеты булавоусых чешуекрылых проводились с 1990-1998 гг. в Юго-Восточном Алтае в хребтах: Сайлюгем, Чихачева, Курайском, Южно-Чуйском, Северо-Чуйском, а также на плоскогорье Укок, в Курайской, Чуйской и Джулукульской котловинах (рис. 1). С учетом сложной орографии Юго-Восточного Алтая и связанной с ней мозаичностью растительности на исследуемой территории были намечены 8 ключевых участков так, чтобы охватить учетами максимальное количество местообитаний.
Первый участок - бассейн р. Тархата (2300-2500 м): от Тархатинского озера до выхода реки в Чуйскую степь, в том числе ее правый приток Каланегир на высотах 2300-2500 м. Учетами охвачены ландшафты степного, лесного, субальпийского и альпийско-тундрового поясов, расположенных в хребтах Сайлюгем и Жумалыкыр. Здесь обследовано 15 местообитаний.
Степной пояс: мелкодерновинно-злаковые-полынные, злаково-осочковые,осочково-злаковые степи, остепненные луга на крутых склонах южных экспозиций. Лесной пояс: парковые лиственничные, ерниково-лиственничные (фрагментарные) леса, подгольцовые лиственничные, ерниково-лиственничные редколесья. Субальпийский пояс: ерники, ерниковые тундры с фрагментами субальпийских лугов. Альпийско-тундровый пояс: каменистые осыпи, каменистые, каменистые-дриадово-лишайниковые, кустарниковые, мохово-травянистые тундры.
Второй участок - бассейн р. Узунтытыгем (урочище Узунтытыгем) охватывает восточную оконечность Курайского хребта на высотах 2500-2800 м. Учеты проведены в 9 основных местообитаниях. Степной пояс: мелкодерновинные полынно-злаково-осочковые степи на склонах южных экспозиций, мелкодерновинные злаково-полынные степи, осочково-разнотравные и остепненные луга на крутых южных склонах. Лесной пояс: парковые лиственничные леса (фрагментарные), лиственничные редколесья. Субальпийский пояс: ерники с фрагментами субальпийских лугов. Альпийско-тундровой пояс: каменистые осыпи южных склонов, щебнисто-лишайниковые, каменистые тундры.
Третий участок - северо-западный макросклон Курайского хребта. Обследование проведено в окрестностях Акташского рудника, урочищ: Баратал и Старые Мёны на высотах в 1500-2900 м. Учеты проведены в 11 местообитаниях.Степной пояс: опустыненно-солончаковые степи Курайской котловины, разнотравно-остепненные склоны, разнотравные увлажненные и остепненные луга на крутых южных склонах. Лесной пояс: лиственничные, кедрово-лиственничные, пойменные лиственнично-кедровые, кедрово-лиственничные редколесья с фрагментами ерников. Субальпийский пояс: ерники, субальпийские луга.Альпийско-тундровый пояс: щебнисто-лишайниковые, каменистые тундры.
Четвертый участок - Джулукульская котловина. Исследования проведены в бассейне р. Богояш и прилегающих склонах хребта Чихачева на высоте 2200-2800 м. В трех поясах обследовано 8 местообитаний. Степной пояс: заболоченные низинные пойменные луга, увлажненные осочко-злаковые луга, мелкодерновинно-злаковые степи. Субальпийский пояс: пойменные ерники, ерники на склонах гор. Альпийско-тундровый пояс: лишайниково-ерниковые, мохово-лишайниковые, щебнисто-лишайниковые и каменистые тундры.
Пятый участок - бассейн р. Себыстей расположен на северо-восточных отрогах Южно-Чуйского хребта. Пять обследованных местообитаний находятся в степном поясе на высоте 2200-2250 м: пойменные заболоченные осоковые закустаренные луга, опустыненные, мелкодерновинно-злаковые, дерновинно-злаковые каменистые степи. Учетом также охвачена юго-западная часть Чуйской котловины с расположенной в ней опустыненной степью.
Шестой участок - бассейн р. Калгуты (левобережье р. Жумалы) по юго-западным отрогам хребта Сайлюгем на высотах 2300-2650 м н.у.м. Учеты проведены в трех местообитаниях степного, субальпийского и альпийско-тундрового поясов соответственно: остепненные варианты южных экспозиций с выходами коренных пород с фрагментарными ерниками и можжевельниками, пойменные травянистые ерники, щебнистые закустаренные тундры.
Седьмой участок обследовался по хребту Чихачева в бассейне р. Бугузун на высоте 2100-2800 м. Учеты проведены в 9 основных местообитаниях. Степной пояс: галечноковыльные тонконоговые и холоднополынные степи юго-восточной окраины Чуйской котловины. Лесной пояс: долинные лиственные редколесья с фрагментами луговых сообществ у нижних граний пояса, лиственичный лес с вырубками, в верхней части хребта лиственнично-кедровые редколесья. Субальпийский пояс: южные каменистые склоны с фрагментами колючеподушечников, закустаренные каменистые осыпи. Альпийско-тундровый: травянисто-ерниковые, травянисто-кустарниковые с каменистыми участками, мохово-лишайниковые тундры.
Восьмой участок - северо-восточные отроги Северо-Чуйского хребта. В пойме р.Чуя (1424-1500 м) учеты проведены: на остепненных лугах, расположенных по крутым склонам южных экспозиций и в опустыненно-солончаковой степи Курайской котловины. На плато Ештыкёль (урочище Ештыкёль) (1800 м) в лесном поясе обследованы: ерники с лиственичными редколесьями. По южным макросклонам Южно-Чуйского хребта, обрамляющих северную часть плато Укок на высоте 2100 м, по левобережью р. Джазатор (могильник Чигур) в степном поясе обследованы: высокогорные мелкодерновинно-злаковые степи. Кроме перечисленных выше местообитаний, в работе использованы опубликованные данные (Бондаренко, Малков и др., 1999) в бассейне р. Джазатор (степь Самаха) на высотах 1600-1700 м. Учеты проведены в 4 местообитаниях степного и лесного поясов. В первом: низинные заболоченные луга в пойме р. Беляши, мелкодерновинно-злаковые участки степи Самаха. Во втором: долинные елово-лиственичные, парковые лиственничные леса. По северо-восточным отрогам хребта Сайлюгем р. Уландрык (урочище Уландрык) (2300-2600 м) учеты проведены в двух местообитаниях: мелкодерновинно-злаково-щебнистые степи, мятликовые степи с фрагментами тундры. Также учетами охвачена Курайская котловина (1500 м), окруженная отрогами Курайского и Северно-Чуйского хребтов: Cтепной пояс: опустыненные степи Курайской котловины - в урочище Моштуарык, разнотравно-злаковые луга- в урочище Тадила. Лесной пояс: пойменные еловые леса- в пойме р.Чуи северо-западнее с. Курай, парковые лиственные леса - у подножия Северо-Чуйского хребта. Субальпийский пояс: ерники на Курайском хребте. По юго-западным отрогам хребта Сайлюгем в бассейне р. Калгуты 2400-2500 м. Степной пояс: злаково-травянистые, мятликово-разнотравные степи. Альпийско-тундровый пояс: травянистые тундры.
Методика учетов описана ранее [2]. Учеты проведены в течение 350 часов на маршрутах протяженностью 875 км. Зарегистрировано более 16500 особей, из 6 семействам, 66 родов и 110 видов. Всего в широком градиенте природно-ландшафтных условий обследовано 79 вариантов комплексов дневных чешуекрылых. Растительные пояса приведены по Н.Г. Огуревой [13].
Для выявления особенностей пространственной организации использовались оценки сходства населения и корреляции их с неоднородностью среды [18]. В результате получено 17 средних вариантов населения дневных бабочек.
Для количественных признаков рассчитаны индексы Жаккара [12], и на их основе проведена факторная классификация с расчетом внутри - и межклассового сходства населения и доразбивкой крупных классов [20].
Материалы математически обработаны с помощью программного обеспечения банка данных лаборатории зоологического мониторинга ИСиЭЖ СО РАН.
Результаты
Изменения плотности и лидеров населения
Общее видовое и фоновое богатство наиболее велико в лесном поясе. Оно уменьшается с высотой, достигая минимальных значений в субальпийском поясе и несколько увеличиваясь в альпийско-тундровом (табл. 1). Плотность населения максимальна в степном поясе и уменьшается как вниз так и вверх от этого уровня, но также, как видовое богатство, несколько возрастает в альпийско-тундровых ландшафтах.
Boloria napaea Hffmsg. и Erebia callias W.H.Edw. лидируют от альпийско-тундрового пояса до степного, хотя в альпийско-тундровом и субальпийском поясах Вoloria napaea Hffmsg. занимает первое место, а в степях - третье. Доля Erebia callias W.H.Edw. с уменьшением высот возрастает до степного пояса, при этом она перемещается в списке лидеров последовательно с третьего на первое место. Остальные виды как лидеры специфичны. В лесном поясе специфичны два из трех лидеров, а один - Lycaeides argyrognomon Brgstr. - общий со степями, где представленность его выше, чем в лесах. В процентном отношении доля Boloria napaea Hffmsg. выше в субальпийском поясе, а Erebia callias W.H.Edw. - в степном и уменьшается вверх и вниз от этих уровней.
В общем следует отметить, что изменения в составе лидеров, как правило, постепенны: вид занимающий первое место, смещается на второе и третье, в то время как виды, находившиеся на этих позициях, выходят на первые места.
| Видов | Плотность, | Доля, | |||
| Пояс | Всего | Фоновых | особей/га | Лидер | % |
| Альпийско-тундровый | 74 | 10 | 38 | Boloria napaea Gr.-Gr. Erebia stubbendorfii Men. Erebia callias Edw. |
16 13 13 |
| Субаль-пийский | 42 | 8 | 30 | Boloria napaea Gr.-Gr. Erebia callias Edw. Mellicta athalia Rott. |
20 17 10 |
| Степной | 89 | 8 | 52 | Erebia callias Edw. Lycaeides argyrognomon Bergst. Boloria napaea Gr.-Gr. |
31 15 8 |
| Лесной | 96 | 11 | 46 | Cupido minimus Fuessl. Lycaeides argyrognomon Bergst. Сyaniris semiargus Rott. |
13 9 9 |
Классификация населения
В населении булавоусых чешуекрылых Юго-Восточного Алтая выделяются три типа сообществ.
1. Сообщества открытых и полуоткрытых необедненных местообитаний - тундр, степей и подгольцовых редколесий (кроме тундро-степей, ковыльных опустыненных и полынных степей), лидеры по обилию - Erebia callias W.H.Edw. и Lycaeides argyrognomon Brgstr. по 15 %; Boloria napaea Hffmsg. 8 %; всего 48 особей/ га, 104 вида, из них 11 фоновых).
Этот тип населения включает 4 подтипа:
1.1 обитатели курумов и крупнообломочных осыпей высокогорий (Erebia pandrose Brkh. 23 %, E. kefershteinii Ev. и E. callias W.H.Edw. по 12 %; 26, 37 - 9);
1.2 -тундр, подгольцовых редколесий, высокогорных разнотравно-злаковых и мелкодерновинных сухих степей (Boloria napaea Hffmsg. 16%, Erebia callias W.H.Edw. 14 %, Proclossiana eunomia Esp. 10 %; 44, 100 - 9);
1.3 - настоящих, разнотравных, кустарниковых каменистых и опустыненных полукустарничковых степей высокогорий (Erebia callias W.H.Edw. 36 %, Lycaeides argyrognomon Brgstr. 15 %, Boloria napaea Hffmsg. 8 %; 66, 60 - 11);
1.4 -лиственичных лесов на границе высокогорий и среднегорий (Proclossiana eunomia Esp.- 36 %,Clossiana freija Thunb. и Melitaea baicalensis Brem. по 8 %; 63, 33 - 14).
2.Сообщества необедненных лесных местообитаний - лиственничных лесов среднегорий (Cupido minimus Fuessl. 21%, Cyaniris semiargus Rott. 15 %, Coenonympha hero L. 8 %; 52, 65 - 10).
1.Сообщества обедненных местообитаний - темнохвойных островных лесов, тундростепей, ковыльных опустыненных и полынных степей (Lycaeides argyrognomon Brgstr.
35 %, Triphysa albovenosa Ersch. 12 %, Polyommatus icarus Rott. 6 %; 17, 46 - 3).
В первом и втором типах плотность населения практически равна, в третьем - в 3 раза меньше. Общее видовое богатство в первом типе в 1,5 - 2 раза выше, чем в двух остальных. Число фоновых видов фактически одинаково в двух первых типах и в 3 раза меньше в третьем. В первом и третьем типах среди лидеров есть общий вид - Lycaeides argyrognomon Brgstr. (15 и 35 %, соответственно). В целом, среди лидеров в первом типе сообществ преобладают тундрово-альпийский и эврибионтный виды, тогда как лесной уступает им почти вдвое. Во втором типе преобладают лесные виды, им в 2 раза уступает эврибионтный; в третьем по убыванию количества распределяются лесной, лугово-степной и эврибионтный виды.
Население необедненных открытых и полуоткрытых местообитаний (1-ый тип) довольно разнообразно, в отличие от двух последующих, и различается предпочитаемыми местообитаниями, в соответствии с которыми в его рамках выделено 4 подтипа. Максимальная плотность населения - в подтипах 1.3 и 1.4, значительно меньшая 1.2 и минимальная в 1.1 (66, 63, 44 и 26 особей/га, соответственно). Общее видовое богатство максимально в подтипе 1.2. Оно почти в 2-3 раза выше, чем в остальных подтипах (100, 60, 37 и 33 вида). По числу фоновых видов на первое место выходит подтип 1.4, несколько меньше их в 1.3 и одинаковое количество в 1.2 и 1.1 (14, 11, 9, 9 видов).
Структурный граф сходства населения дневных бабочек Юго-Восточного Алтая построен при пороге значимости выше 11 единиц (рис. 2).
На нем видны сгущения подтипов (1.1-1.4), относящихся к одному типу. Mежподтиповое сходство населения, как правило, велико (20-21 единица). Для сообществ лиственничных лесов среднегорий (второй тип) и обедненных вариантов третьего типа оно меньше - 11-13. На рисунке показана ориентация структурного графа в двух- (рис. 2 А) и трехмерном (рис. 2 Б) пространстве (если учесть следующую по значимости связь междунаселением подтипа 1.3 и типа 3). На схеме видны два основных направления изменений населения. Первое связано с теплообеспеченностью, определяемой абсолютной высотой местности. В том же направлении изменяется скальность и облесенность. Отклонения от основного тренда определяются остепнением (1.3), независимым влиянием инсуляризации, опустыниванием и продуктивностью (3).
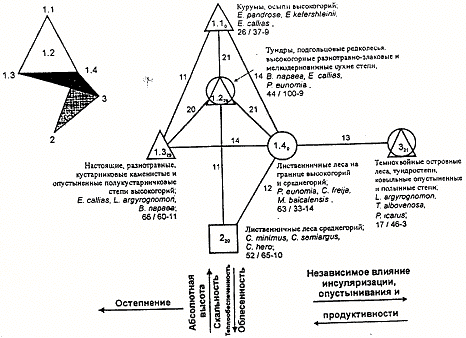
Рис. 2. Пространственно-типологическая структура населения булавоусых чешуекрылых Юго-Восточного Алтая: а - двухмерное пространство, (треугольниками обозначены открытые местообитания, кружками - полуоткрытые, квадратом - лесные местообитания). б - трехмерное пространство.
Максимальная плотность населения и богатство фоновых видов свойственны настоящим, разнотравным, кустарниковым каменистым и опустыненным полукустарничковым степям высокогорий (66 особей/га и 11 фоновых видов). Почти столько же бабочек (63) при большем числе фоновых видов (14), зарегистрировано в лиственничных лесах на границе высокогорий и среднегорий. С увеличением абсолютных высот уменьшается теплообеспеченность, и, как правило, продуктивность растительности, что приводит к уменьшению суммарного обилия и видового богатства булавоусых чешуекрылых. Кроме того, неоднородность населения коррелирует, хотя и в меньшей степени, с остепнением, облесенностью и затененностью в лесах, что приводит к уменьшению общего числа встреченных видов. Из выявленных структурообразующих факторов наиболее значима теплообеспеченность, определяемая абсолютными высотами, и в меньшей степени - увлажнение, поясность и облесенность (табл. 2).
Множественная оценка силы и общности связи неоднородности населения и структурообразующих факторов среды за счет их скоррелированности учитывает 46 % дисперсии населения, отраженной матрицей коэффициентов сходства. Информативность классификации населения составляет 65 %, выявленной структуры - 71 % дисперсии. Общие представления о факторах и природных режимах по классификации и структуре населения при значительной их корреляции совпадают с неоднородностью среднелетнего населения дневных бабочек на 83 %.
| Фактор, режим | Учтенная дисперсия, % |
| Теплообеспеченность (абсолютные высоты) | 25 |
| Увлажнение | 10 |
| Поясность | 9 |
| Облесенность | 8 |
| Скальность | 6 |
| Всего факторы | 46 |
| Всего режимы по классификации | 65 |
| Всего режимы по структуре | 71 |
| Всего факторы и режимы | 83 |
Заключение
Общее видовое и фоновое богатство населения булавоусых чешуекрылых максимально в лесном поясе и в целом уменьшается с высотой, хотя в альпийско-тундровом поясе несколько увеличивается. Плотность населения наиболее высока в степном поясе и уменьшается вниз и вверх от этого уровня. Основные изменения в населении связаны в Юго-Восточном Алтае с теплообеспеченностью территории, которая в свою очередь определяется абсолютной высотой местности, а также с поясностью и облесенностью.
Исследования, послужившие основой данной статьи, поддержаны Министерством образования Российской Федерации <Конкурс 2002 года на соискание грантов на проведение молодыми учеными научных исследований в ведущих научно-педагогических коллективах> и грантом Президента Российской Федерации c МК - 2500.2003.04 для поддержки молодых российских ученых и ведущих научных школ Российской Федерации>.
 2. - С. 121-129.
2. - С. 121-129.